
Library
Browse resources published by our research team.
In addition to full texts of our peer-reviewed articles, our library includes research digests that break down our peer-reviewed articles; in-depth reports that thoroughly examine a topic; commentaries that explain the significance of particular issues in wild animal welfare science; and short communications that briefly survey a field or topic.
Wild Animal Initiative adheres to Open Science TOP Guidelines. Read more here.
Quantifying the neglectedness of wild animal welfare
Michaël Beaulieu's deep dive examines the quality and quantity of mentions of wild animal welfare in the scientific literature and finds it to be a neglected research area.

Questioning the neglectedness of wild animal welfare: passionate claims, illusionary facts, or simple truth?
Wild animal welfare is often suggested to be a neglected research area, worthy of more attention from scientists and policy makers (e.g., Tomasik 2015; Forristal 2022). Considering the general propensity of animal welfare to unleash passions even among scientists (Coghlan & Cardilini 2022), it is important to assess the veracity of these claims. So to objectively assert the need to conduct more research to better understand wild animal welfare and implement sound welfare interventions in the wild, clear evidence (or “stubborn facts” insensitive to people’s inclinations and passions; Adams 1770) supporting the neglectedness of wild animal welfare is required.
Cursory research of the expression “wild animal welfare” in Google Scholar (Nov. 2023) provides 844 outputs, only 7% of which emphasize the expression in their title or abstract. However, such figures may be misleading if wild animal welfare is already considered under alternative synonyms or already incorporated into adjacent disciplines. For instance, the fate of wild animals in response to variable environmental conditions has traditionally been under the scrutiny of conservation practitioners (Brown 2007; Nijhuis 2020). Because conservation studies typically focus on conditions that affect animals in their natural habitat (for instance, environmental conditions changing because of anthropogenic activities and that may be perceived as unpleasant by wild animals), they may theoretically also encompass aspects related to wild animal welfare (Sekar & Shiller 2020). Therefore it is possible wild animal welfare is not so highly neglected, but rather often concealed within existing animal conservation studies. Importantly, because the multidisciplinary field of wild animal welfare includes a variety of approaches, indicators, and taxa, some research areas within this field may be more neglected than others.
Assessing the neglectedness of wild animal welfare
To assess the neglectedness of wild animal welfare, I have conducted a targeted review of the literature in peer-reviewed journals publishing research in animal welfare (three journals: Animal Welfare, Journal of Applied Animal Welfare Science, animal welfare section of Animals) and animal conservation (five journals: Animal Conservation, Biological Conservation, Conservation Biology, Conservation Physiology, Oryx). I selected these journals because of their high visibility and their representativeness in their respective field. I considered a date range of 2013-2022 for the review process, in order to reflect contemporary research on animals. Accordingly, I only included research articles (as opposed to reviews, commentaries) focusing on animals (as opposed to, e.g., plants, policy) in the review of both fields. I identified and classified the study subjects of each welfare science article as “wild” if they were studied in their natural habitat or collected in the wild and then studied in captivity. In conservation articles, I searched for welfare-related terms (e.g., welfare/wellbeing, sentience, emotion, conscious, subjective experience, affective state, compassionate) in both the title and abstract.
Confirming the neglectedness of wild animal welfare
Both the presence of wild animals among all reviewed welfare studies (6%) and the occurrence of welfare-related terms in conservation studies (1%) were very low (Fig. 1). Among welfare-related terms, “welfare/wellbeing” was by far the most commonly used term (90%) in conservation journals. In addition to “welfare/wellbeing,” other welfare-related terms such as “compassionate” (6%) and “sentience” (4%) were also found. However, other welfare-relevant terms such as “conscious,” “subjective experience,” or “affective state” were completely absent. Altogether, these proportions strongly suggest that wild animal welfare is neglected by both the welfare and conservation literature.

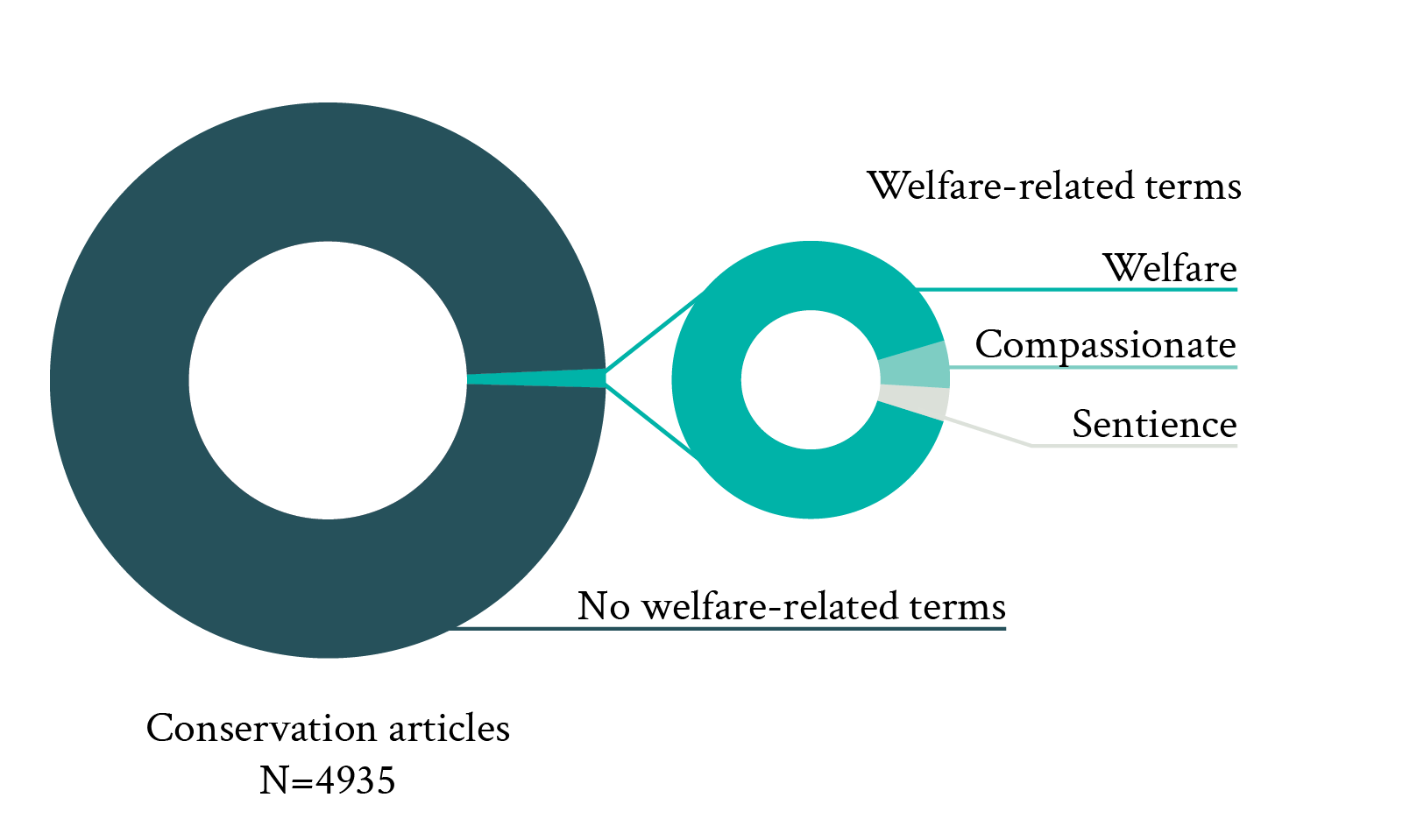
Figure 1. Proportion of articles published in three principal animal welfare journals between 2013 and 2022 and including wild animals (left pie, top on mobile), and proportion of articles published in five principle conservation journals over the same period and including welfare-related terms in either their title or abstract (welfare, compassionate and sentience were the only terms found; right panel, bottom on mobile).
Dissecting the neglectedness of wild animal welfare: the example of physiological approaches
To understand how an animal experiences their life in the wild, it is necessary to use approaches that can evaluate and provide evidence of their welfare. Behavioral and physiological markers potentially represent downstream indicators of welfare, and can thus provide insight into the welfare of wild animals (Browning 2022). I focused here on physiological markers specifically, as they may be highly informative (on their own or to complement behavioral approaches) for assessing the welfare of wild animals (see deep dive: Welfare and physiology: a complicated relationship). To quantify the degree to which physiological approaches are also being neglected in wild animal welfare science, I have further examined the welfare science studies previously identified as being conducted on wild animals to determine whether they made use of physiological markers and in which species.
Only 22% of welfare science studies conducted on wild animals included the use of physiological markers (Fig. 2). These results suggest that wildlife biologists either are reluctant to make use of physiological markers in their research relative to other welfare indicators (e.g., living conditions, behavior), or they lack awareness of how to incorporate them. Yet the very existence of the field of conservation physiology, and the numerous studies published, demonstrate that it is possible to measure a diversity of physiological markers in the wild to examine how wild animals are affected by given environmental conditions (Cooke et al. 2013). Nevertheless, despite this potential, thus far such studies have not focused on assessing welfare, as indicated by only 3% of articles published in conservation journals and including physiological parameters mentioning welfare-related terms in their title or abstract. Moreover, the few studies that considered physiological parameters to assess welfare in wild animals focused on a limited number of physiological markers (compared to the vast variety of physiological parameters that could theoretically be measured to assess welfare; e.g., Jerez-Cepa & Ruiz-Jarabo 2021; Whitehead & Dunphy 2022) measured in few taxa, the typical study measuring glucocorticoids in a mammal species (Fig. 2).
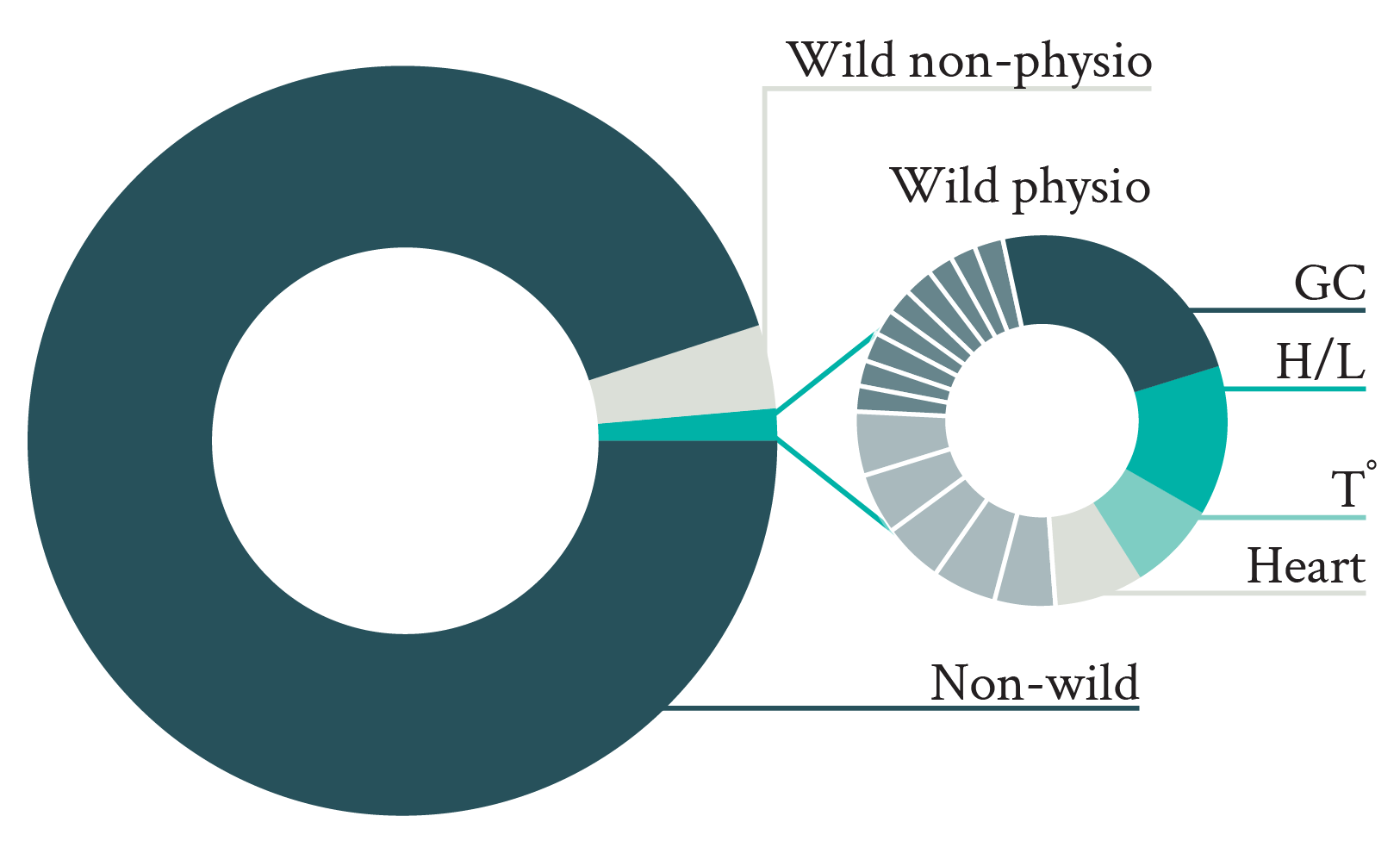
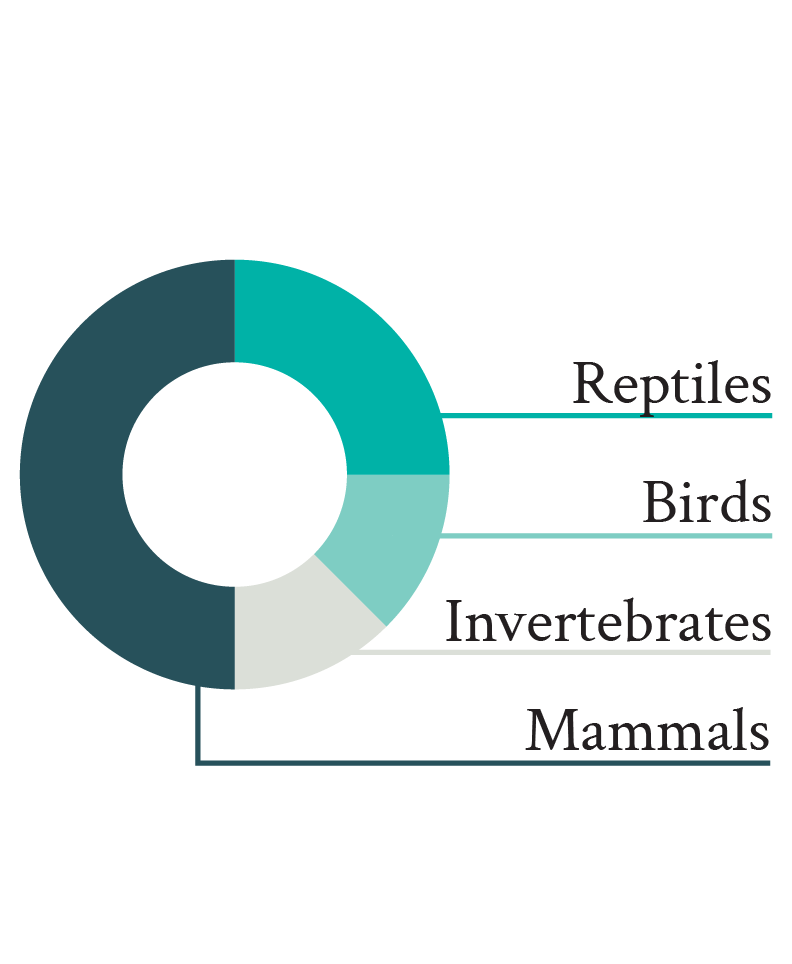
Figure 2. Proportion of articles published in three principal animal welfare journals between 2013 and 2022 and including wild animals (left pie, top on mobile) and measuring physiological markers (striped area). The detail of the measured physiological markers (GC: glucocorticoids; H/L: heterophil/lymphocyte ratio, Tº: temperature) and in which taxa (Invert.: invertebrates) they were measured is illustrated in the right pies (bottom on mobile). For clarity, only the first half of the physiological markers measured in articles on wild animal welfare physiology is annotated.
The overrepresentation of glucocorticoids (and their effects on the H/L ratio; Davis et al. 2008) and mammals among studies on the physiology of wild animals is not necessarily surprising, but their measurement is questionable when examining their welfare. Indeed, even though measuring glucocorticoids reflects a general trend observed in animal stress physiology (MacDougall-Shackleton et al. 2019), the relationship between glucocorticoids and welfare remains unclear (Ralph 2016). Therefore, for glucocorticoid measurements to offer useful insights about welfare, they need to be complemented by additional markers (e.g., behavioral observations). Moreover, even though the imbalance in favor of mammals probably reflects the widespread greater interest in these animals, mammals are relatively rare in nature (ca. 0.1% of all animal species and ca. 0.35% of the whole animal biomass on Earth; Mora et al. 2011; Burgin et al. 2018; Bar-On et al. 2018), and the way they experience their lives is probably not comparable to that of other taxa with very different neural organizations (e.g., more common invertebrates; Paul et al. 2020).
Conclusions
Review of recent literature in key peer-reviewed journals in animal welfare showed that the neglectedness of wild animal welfare is indeed a “stubborn fact” and not just a questionable claim. Importantly, the targeted review of journals in animal conservation also provided evidence that this underrepresentation was not simply an artifact of the inclusion of welfare aspects within animal conservation studies (that were found to refer to animal welfare only anecdotally and superficially). Moreover, within the limited number of studies that did focus on wild animal welfare, the limited use of physiological markers further highlights the neglectedness of valuable methodological approaches and taxa. If allowed to persist, such biases will likely limit our understanding of wild animal welfare, as the strength of this field precisely lies in its diversity of approaches and the range of taxa to which they can be applied. Continuing not to take advantage of such potential would be short-sighted.
The limited availability of welfare science studies focusing on wild animals suggests that there may be resistance or barriers to extending the scope of welfare science beyond the traditional context of farmed, companion, and captive-housed animals. A contributing factor may be that assessing welfare is easier in well-known model species maintained under controlled conditions in captivity (e.g., pigs), compared with their wild counterparts living under uncontrollable conditions (e.g., wild boars). Another contributing factor might also be related to the uncertainty about the capacity of many wild animals to have subjective experiences (e.g., invertebrates, but see Anderson & Adolphs 2014; de Waal & Andrews 2022). Despite these inherent challenges, there is increasing evidence that extending the field of animal welfare to wild conditions is feasible (Harvey et al. 2020), particularly through collaborations among scientists from a range of disciplines, and could have far-reaching impacts for our general understanding of how most animals on Earth experience their lives. In addition to advancing general understanding and increasing available knowledge, wild animal welfare studies may also lead to concrete applications — for instance, by informing stakeholders, refining practices, or modifying legislation impacting wildlife. But the full potential of the field of wild animal welfare will be reached only if scientists branch out beyond traditional norms and consider the full range of approaches and taxa that this field has to offer.
Biotelemetry provides insight into a wild eagle’s death
Jane Capozzelli
Jane Capozzelli reviews how biologgers can provide information about welfare, but also pose risks to animals, following the case of a single eagle.
Key takeaways
- Biotelemetry can provide a high level of detail about wild animals’ activities.
- Real-time behavioral and physiological data can indicate how an animal might be feeling.
- Location data can link welfare indicators to the animal’s environmental context.
- Although biotelemetry could advance efforts to improve wild animal welfare, its methods can be harmful or lethal to the animals being studied.
- Wild Animal Initiative will continue to evaluate the costs and benefits of biotelemetry. We hope to identify opportunities to collect highly detailed individual-level data without burdening the animals involved.
Introduction
Biotelemetry devices (“biologgers”) can monitor behavioral and physiological indicators of welfare status in real time, and couple these data with precise information about environmental context (Brown et al. 2013). Biologgers consist of a suite of sensors attached to an animal. The different sensors typically record location, motion, or other biological and environmental features multiple times a day (Cooke et al. 2004). The data collection is remote, meaning that the human observer does not operate in the field except to capture the animals and attach the devices.
In one such study, ornithologists outfitted a female white-tailed sea eagle (Haliaeetus albicilla) with a biologger that recorded her movement, circadian rhythm, behavioral profile, and body temperature (Krone et al. 2009). In this article, I use this case study to illustrate biotelemetry’s potential to offer insights on wild animals’ welfare.
The biologger in this case comprised two radio transmitters, a battery pack, and sensors monitoring location, temperature, vertical motion, and horizontal motion. (Figure 1). At 170 g, the device weighed 3.5% as much as the eagle. The study ended after only five months when the eagle unexpectedly died.

B.

Figure 1
A) The biologger attached to the white-tailed sea eagle (Haliaeetus albicilla). Adapted from Krone et al. (2009). B) An eagle outfitted with a biologger comparable to the “backpack” style used in the sea eagle case study. Photo credit: Alan Schumacher/USDA Wildlife Services.
As with many biotelemetry studies, this one focused on a rare species (the white-tailed sea eagle) and collected data to inform its conservation. Although welfare was not the focus of the project, it nonetheless proved data highly relevant to individual welfare because it documented the eagle’s dying process in unprecedented detail. This illustrates how biologgers could be used to understand the welfare burdens of different causes of death.
Welfare-relevant conclusions from biotelemetry
Cause of death
GPS data enabled the researchers to identify both the immediate cause of death and the broader environmental context for it.
Because the biologger transmitted location data every 5 minutes, the researchers were able to determine the precise time and location of the eagle’s death and to recover her corpse as soon as possible — overcoming major challenges in cause-of-death research. Through necropsy, they determined she died of lead poisoning (Krone et al. 2009).
Because the data stream was continuous for 5 months, there was strong evidence that the eagle had ingested lead while foraging within her territory. Even though the eagle had a high-quality habitat that was protected in a conservation reserve, she was still at risk of lead exposure because of legal hunting in her environment (Krone et al. 2009).
Welfare while dying
The biologger fitted to this eagle also included accelerometers, which provided a high-resolution behavioral profile. Activity readings were taken 8 times per second for the entire duration of the study. This frequency enabled researchers to characterize the eagle’s usual circadian rhythm, as well as the start of her sickness period and her dying period. The sickness and dying periods each lasted 11 days (Krone et al. 2009).
With an understanding of the animal’s typical daily rhythms, it was clear when she started showing abnormal behaviors that indicated disease: changes to the timing of flight and rest and increases in the frequency of rest periods (Veissier et al. 1989, Scheibe et al. 1999). The eagle eventually ceased all activity. This extreme lethargy signalled the beginning of her dying process (Krone et al. 2009). Her body temperature also dropped a total of 15℃, coinciding with the decreases in activity (Krone et al. 2009). This physiological and behavioral profile provides a basis for inferring welfare status during the dying process (Figure 2).

Figure 2
Using biotelemetry data to estimate welfare. The activity and body temperature timelines approximate the actual data Krone et al. (2009) collected from a biologger attached to a white-tailed sea eagle (Haliaeetus albicilla) in the weeks preceding her death. The inferred feelings timeline estimates the subjective mental states the eagle may have been experiencing at the same time.
I compared the eagle’s behavioural and physiological profiles over time to her baseline readings in order to estimate her mental state (Beausoleil et al. 2018). Although it may be hard to ever know with certainty how physiological conditions relate to subjective feelings in other species, we can make reasonable guesses given similarities between our own and other animals’ needs.
The eagle’s lethargy could be associated with negative feelings of malaise. The sickness and dying phases spanned 22 days, so the eagle would have also had increasing feelings of hunger, as she was not eating during this time (Mellor and Beausoliel 2015). The necropsy showed total depletion of fat reserves (Krone et al. 2009). Since eagles eagles ideally eat about 5% of their body weight daily to maintain peak body condition, (Fevold and Craighead 1958), it is reasonable to assume that this loss of reserves and lack of hunting activity would lead to feelings of severe hunger and discomfort. Lastly, the 15º C drop in body temperature would likely lead to extreme discomfort associated with coldness (Mellor and Beausoliel 2015).
Advantages of biologgers for wild animal welfare studies
Recovering corpses
For studying death in the wild, the traditional methods such as short-duration video or corpse-recovery surveys have well-known limitations. Corpses are encountered by chance (Dell et al. 2014). Most are not recovered because they are decomposed or scavenged, making it incredibly difficult to estimate the relative frequencies of different causes of death in a population (Tavecchia et al. 2011).
Biotelemetry has major advantages for cause-of-death research because it provides detailed information on when and where animals die (Klaassen et al. 2014). As a result, most of the deaths can be attributed to a specific cause (Figure 3). The positional data can also help researchers rapidly and systematically recover corpses (Calvete et al. 2002).

Figure 3
Causes of death in eagles who died during biotelemetry studies. Data from a cause-specific mortality database (Hill et al. 2019a).
Knowing the prevalence and severity of different causes of death is important, because the amount of suffering that an animal experiences while dying is largely determined by the cause and manner of death (Beausoleil et al. 2016). Vehicle collisions, hunting with firearms, or electrocution may cause near-instantaneous death, whereas disease, starvation, or intoxication may weaken the animal over an extended period (22 days in our eagle case study).
Beyond simply identifying the cause of death, comparative studies could allow us to infer the experiences of animals that die from different causes, and how the prevalence of these causes varies by life stage or environment. Such knowledge would allow wild animal welfare advocates to prioritize preventing the most painful manners of death.
Observing behavior
Currently, most welfare assessments are done by in-person human observation (Bailey et al. 2018). But biotelemetry allows continuous data collection from almost any location an animal may occupy — even where people cannot follow (Ropert-Coudert and Wilson 2005).
The ability to record quantitative behavioral profiles anywhere the animal goes allows for much more informed welfare estimates. For example, behavioral profiles can show sickness behaviors or other abnormal trends in function and condition (Brown et al. 2013, Adelman et al. 2014). This multidimensional information allows us to understand an animal’s welfare status in the context of events and environmental variables that matter at the scale of an individual’s preferences and perceptions (Bisson et al. 2009, Martins et al. 2014, McClune et al. 2015, Clemente al. 2016).
Observing community interactions
Because biologgers minimally distort behavior and can transmit data from any location, they are particularly well-suited to observing individuals’ interactions with their communities. For example, animals in different environments could be monitored as they cope with the various habitats that they encounter. This analysis would provide an animal’s-eye view on habitat quality and show to what extent an animal’s welfare might be improved or diminished by altering the environment (Lima and Zollner 1996, Nicol et al. 2020).
By pairing biotelemetry with an understanding of community interactions, it may be possible to show how community structure degrades or enhances the average welfare of each animal community-member. For example, in a community with two species that cooperate beneficially, biotelemetry could monitor the degree to which stress is reduced when animals have the opportunity to interact (Hammers et al. 2019).
Concerns about biotelemetry research
Technological limitations
Cost is the largest barrier to scaling wildlife biotelemetry research. Devices typically range from hundreds to thousands of dollars per unit (Cooke et al. 2004). Because of the high cost, sample sizes of biotelemetry studies are typically insufficient to predict a welfare outcome before it occurs, even though the data itself is reflective of welfare status (Lindberg and Walker 2007, Costantini and Møller 2013, Naef-Daenzer and Grüebler 2014).
Additionally, while biologgers have been shrinking in size due to advances in battery technology, they are still most commonly only usable on larger animals (Cooke et al. 2004). Unless the weight barriers can be overcome, the vast majority of animals — small invertebrates — are impossible to study with biotelemetry.
Among marine invertebrates, biotelemetry is generally only feasible for individuals weighing more than 400 g. If this threshold were decreased to 40 g, then ten times more species could be studied, as could larvae and juveniles (O’Dor 2002). In terrestrial insects, biotelemetry studies have been limited to a few large arthropods, such as locusts or hawkmoths. Insect biologgers usually weigh less than 1 g, but their battery life is 3 hours or less (Sato and Maharbiz 2010). This temporal limitation nearly eliminates the ability to research invertebrate welfare through biologgers.
Welfare harms
Biologgers harm the animals wearing them (Hawkins 2004). Death during the tagging process can occur either at capture, directly after release, or from physical attrition over the course of the study (Casas et al. 2015, Weiser et al. 2016, Lameris et al. 2018). For example, the eagle in our case study was caught with a noose, a particularly stressful and risky method for capturing birds (Kautz and Seamans et al. 1980).
Even in cases with no tag-related mortality (for example, Goldsmith et al. 2017), the devices can have sublethal impacts such as exhaustion, injury, reduced body condition, and the disruption of social activities and pair bonding (Weiser et al. 2016, Brlík et al. 2020, Hamelin and James 2018, Lameris et al. 2018; but see Niles et al. 2010 which found no sublethal impacts).
The eagle in our case study appeared to have no negative symptoms from the biologger (Krone et al. 2009). However, the authors noted several instances where the bird was at rest during abnormal periods of the day. The authors theorized that stresses such as hunters, tourists, inclement weather, or even sub-lethal lead exposures, may have disrupted the eagle (Krone et al. 2009). But back-mounted biologgers such as the one worn by this eagle (Figure 4) have been documented to cause significant drag that impairs flight (Irvine et al. 2007, Meyer et al. 2015). Therefore, the device itself could have periodically exhausted the bird and necessitated abnormal rest periods.
Conclusions
Biotelemetry’s scientific applications are promising for wild animal welfare research. For example, biologgers can be used to understand the prevalence of different causes of death. Biologgers provide behavioral and physiological profiles at such an extraordinary level of detail that they can potentially be used to infer animals’ mental states. As this study illustrates, these details can be discovered not only throughout an animal’s life, but uniquely, during their sickness and dying process, potentially revealing important information about the harms associated with different causes of death. Finally, by linking physiological and behavioral data with precise location data, biotelemetry gives unprecedented insight into the links between animals and their environment in real time.
The shift from direct human observation to remote monitoring is not without costs to the animals themselves. Biologgers’ weight and drag is substantial, and there are many documented instances of welfare harms and tag-related deaths. Therefore, it remains unclear whether biologgers in their current forms are justifiable or necessary for conducting wild animal welfare research. We plan to conduct further work to identify the costs and benefits associated with biologgers, best practices for determining when such work is necessary (if ever), and what can be done (if anything) to eliminate welfare harms during remote monitoring studies.
How wild animals die: what we know so far
In part three of this series, Luke Hecht gives an overview of what research in this field has taught us so far about how wild animals die and highlights gaps that seem especially important for welfare biology.
This is the final post in three-part series. Read part one → Read part two →
Key takeaways
- Many studies have investigated the causes of death of adult mammals and birds, but there is a lack of data on the deaths of juvenile animals, as well as fish and insects.
- Large, adult animals are more likely to be killed by humans than by any other cause. Small-bodied animals and juveniles are more susceptible to predation by other wild animals.
- Technological improvements in underwater monitoring will improve our understanding of wild fish mortality.
- Insects often die in very different ways from larger animals. Most of what we know about insects’ deaths comes from agricultural pest control research.
- Populations need to be studied under a range of conditions to predict how their cause-specific mortality rates might change under different scenarios.
Introduction
Previously, I wrote about approaches to studying wild animals’ causes of death, with the goal of making work in this field maximally useful for understanding wild animal welfare. Cause of death has not received sufficient research attention relative to its significance to wild animal welfare. The overwhelming majority of the research that does exist is focused on land-dwelling mammals and birds, and primarily on cases where understanding what animals are dying from is instrumental to preventing the extinction of their species. Here, I will give an overview of what research in this field has taught us so far about how wild animals die and highlight gaps that seem especially important for welfare biology.
Cause of death in wild terrestrial vertebrates
Collins and Kays (2011) conducted the first systematic review of cause-specific mortality rates in mammals, selecting only studies that used small radio trackers to monitor animals and document their deaths in a timely manner. They also limited their analysis to adult animals, due to a serious lack of data on juvenile mortality causes. This review found that, overall, predation was the most common cause of death for small-bodied mammals, while human-caused deaths, including hunting and vehicle collision, were the most common causes of death in larger mammals.
In 2019, Hill et al. expanded on this review with a vast amount of new data, including data from juveniles and non-mammalian vertebrates. They found that natural causes of death (predation, disease, starvation), especially predation, were common among juvenile animals irrespective of their species’ typical adult body size. For mammals and reptiles, predation was roughly twice as frequent in juveniles as in adults, with predation accounting for more than 95% of documented deaths of juvenile reptiles (Figure 1). Birds, on the other hand, exhibited very similar cause-specific mortality rates between juvenile and adult age classes.
The authors note that their dataset probably still greatly underestimates natural causes of death, considering that these deaths are often harder to detect. Although smaller animals exhibit the highest natural mortality rates, they are harder to monitor. Research to date has also focused on populations that come into conflict with or are used by humans. These factors combine to make the most numerous deaths in the wild the least understood.

Figure 1
Cause-specific mortality rates separated by age and taxonomic class. Adapted from Hill et al. (2019).
Hill et al. provide an excellent summary of mortality causes, but many of these categories — especially predation, accident and disease — encompass a huge degree of variation in experience. For example, 18% of New Zealand sea lions die from tuberculosis, while 24% die as a result of violence from other sea lions (Lenting et al. 2019). Similarly, approximately half of American black bears die as cubs, often as a result of attacks by older male bears (LeCount 1987). In their study area of northern Pennsylvania, Alt (1984) estimated that 5% of cubs drowned in their winter dens as a result of flash flooding. This was 3-5 times more likely to occur when dens were made in root cavities or excavated soil. Juvenile bears also frequently die from disease (e.g. Chomel et al. 1998). For example, a grizzly cub suffering from canine hepatitis died live on camera in Katmai National Park beside their bewildered mother and siblings. Welfare interventions will need to account for all this variation in individual wild animals’ experiences of death.
Much of the data on birds’ deaths comes from urban wildlife hospitals, where traumatic injury is often the apparent leading cause of death among adult birds, while disease and malnutrition is responsible for the majority of juvenile deaths (e.g. Stenkat et al. 2013). However, these statistics are certain to be biased by the kinds of birds and causes of death that people are most likely to come into contact with. For example, although many traumatic injuries are attributable to domestic cats, birds attacked by predatory raptors are rarely admitted (c.f. Palma et al. 2006). Studies of radio-tagged wild birds report predation and hunting as the leading causes of death (Hill et al. 2019). Some disease mortality can also be linked to hunting practices, as the importation of birds to stock hunting grounds can contribute to outbreaks (Buenestado 2009). Migratory birds present a special challenge for researchers, since many of their deaths occur in transit over long distances, where it may be impossible to recover their corpses. For example, out of 51 probable deaths among satellite-tracked migratory raptors, Klaasen et al. (2014) were only able to confirm 10, among which the leading causes of death were collision with man-made structures and exhaustion during migration.
The progress made in the last decade of cause-specific mortality research is encouraging, but there is much more to be learned, especially about more numerous, smaller-bodied animals. Little is known about the causes of death among prairie dogs, for instance, except that predation is common and plague occasionally wipes out entire colonies (Crosby and Graham 1986; Stapp et al. 2004). Studies of wild reptiles, especially snakes and turtles, consistently indicate road traffic accidents as a leading cause of death (e.g. Himes et al. 2002), although most juveniles are killed by predators (e.g. Butler and Sowell 1996). These gaps in our knowledge of wild animal mortality seem likely to be filled as radio tracking devices are made smaller, lighter and more resilient over the next decade (Kays et al. 2015).
Cause of death in wild fish
Among vertebrates, the most neglected group in the current body of cause-specific mortality research is wild fish. Of the papers that do touch on causes of death in wild fish, many are case reports focusing on the risks of disease outbreaks to human health or the fishing industry. Case reports can be valuable for demonstrating the presence of specific risks that some number of fish do die from (Krkošek 2017). For example, Sterud et al. (2007) detected an outbreak of the parasite Tetracapsuloides bryosalmonae in an Atlantic salmon population that appeared to be their most common cause of death during the study period. This parasite is probably in constant circulation among the fish, and outbreaks have been documented in many other populations, but it is impossible to say whether these results are a representative snapshot of the parasite’s effect on mortality or what alternative causes of death occur in the population.
Most effective studies of terrestrial animals’ cause of death rely on recovering the corpses of tagged individuals, as discussed in my previous post. However, this is clearly more challenging to do with fish. Radio signals for tracking individuals are relatively ineffective underwater, and corpses are much less likely to be recovered (c.f. Benelli and Pozzebon 2013). Several studies have cleverly taken advantage of avian predators naturally retrieving radio-tagged fish from the water and depositing their eaten corpses on shore. For example, Koed et al. (2006) estimated that 39% and 12% of juvenile salmon and brown trout, respectively, were preyed upon by cormorants in the estuary where their study was set. Approximately a further 10% of salmon and trout were killed by cormorants in the adjoining river, while 3-6% were preyed upon by pike, a predatory fish. A similar study by Dieperink et al. (2001) recorded that 65% of juvenile trout were eaten by cormorants and herons while migrating downstream to the Baltic Sea. Fritts and Pearsons (2004) used another predator-based approach to quantify predation, analyzing the stomach contents of smallmouth bass to estimate their annual consumption of salmon. Taken at face value, their results suggested that smallmouth bass accounted for only around 4% of salmon mortality in this system, though the authors noted that this was probably an underestimate. Despite the special challenges involved in recovering the corpses of wild fish, it is at least possible to study the impact of specific predators on fish populations.
Technological progress is proving especially important for studying cause of death in the marine environment. In one recent study, fish were implanted with acoustic transmitters that enabled the researchers to detect predation events occurring within range of their underwater receiver array (Weinz et al. 2020). They inferred that approximately one third of tagged fish were preyed upon over the four months following their release. Several factors could be confounding this result, in both directions. For one, the invasive procedure required to implant the fish with acoustic transmitters could have made these particular fish more susceptible to predation than average. This would be consistent with other studies showing that fish caught and released are more vulnerable due to physical injuries or stress (e.g. Raby et al. 2013). On the other hand, some predation events were likely missed because they occurred out of range of the acoustic array. As the range of underwater tracking improves and the size of transmitting devices decreases so as to cause less harm to the tagged individuals, we should expect to see much more data on the lives and deaths of fish and other marine animals.
Cause of death in insects
Insects are among the most numerous and diverse animals, but also the smallest. Their diversity and size makes them especially difficult to monitor on an individual level. At the same time, some insect species are viewed as pests, and so there is commercial interest in understanding their natural causes of death to devise more effective management strategies (Roux and Baumgärtner 1998). This has motivated a handful of studies reviewing the causes of death of agriculturally relevant insects (Table 1).
Some of these studies use a combination of field observations and population modeling to estimate by how much the population’s overall mortality rate would be reduced if a given cause of death could be eliminated. For example, if predators disproportionately target weak or sick prey, then even if many individuals ultimately die from predation, eliminating their predators may have only a small effect on the overall mortality rate because the same individuals who are vulnerable to predation are also vulnerable to dying soon from disease or starvation.

Table 1
Summarized results from eight studies on cause-specific mortality in six insect species from a pest management context.
Determining the significance of competing mortality risks
Future interventions seeking to improve wild animal welfare should account for how actions to protect animals from specific causes of death could influence population sizes, lifespans, and alternative causes of death. Compensatory mortality, where mortality due to a certain cause is replaced by mortality due to another cause when the first is removed, has been noted in many diverse species and ecosystems. For example, Hostetter et al. (2012) reported that cormorants along the Columbia River in Oregon selectively predate poorer-condition juvenile steelhead salmon en route to the sea. Had they not been killed by cormorants, these same individuals would likely have been among the first to succumb to harsh conditions and competition during their first winter in the ocean (Hurst 2007). Similarly, in a classic study on mule deer, Bartmann et al. (1992) found that removal of coyotes over winter reduced the rate of mule deer deaths by predation, but did not increase their overall survival rates, as the researchers observed increased mortality due to starvation. Diseased mule deer are also selectively preyed upon by mountain lions (e.g. Krumm et al. 2009), which suggests that some of this predator-caused mortality is compensatory.
On the other hand, some cause-specific mortality really is irreplaceable. Achhami et al. (2020) demonstrated this for plant chemical defenses against wheat stem sawflies, while Cooley et al. (2009) showed that other causes of death are not reduced in populations of mountain lions hunted by humans. Bergman et al. (2015) proposed that the degree to which mortality due to a particular cause is compensatory depends on ecological context. For example, if chronic illness made animals less effective at competing for food in a dense population, death by starvation may occur first and compensate for some disease-related mortality in this hypothetical population. If food later became more accessible, we might observe an apparent increase in mortality caused by diseases that are now able to run their course. In addition to studying mortality dynamics in a snapshot of a population’s present conditions, research should model how mortality risks compete under different conditions (Siler 1979) (Figure 2).

Figure 2
Three hypothetical scenarios for the dynamics of mortality due to predation on a prey population. In all scenarios, baseline mortality in the absence of predators is 50 individuals per year. If predator-induced mortality is completely additive (purple diamonds), 10 more animals dying to predation increases total mortality by 10. If predation is completely compensatory (blue squares), total mortality stays constant despite an increased number of deaths by predation; instead, there are commensurately fewer deaths by other causes. In most cases, mortality due to a given cause is likely to be partially compensatory (green circles), lying somewhere between these two extremes. In this model, low rates of predation are mostly compensatory, perhaps removing old or sick individuals from the population. As the intensity of predation increases, a larger proportion of healthy animals are killed and so predation mortality becomes increasingly additive.
Conclusions
A relatively comprehensive snapshot of cause of death in the wild is emerging for terrestrial mammals, birds, and reptiles, thanks to mainstream research in ecology and conservation (Hill et al. 2019). For example, we can see that predation is the leading natural cause of death, especially in juveniles (Figure 1). However, this account is still likely to be heavily biased, since most studies focus on adults and large-bodied species. The focus on large animals could be inflating the number of deaths caused by hunting rather than vehicle collisions, for instance, given that accidents involving small animals often go unreported (Sáenz-de-Santa-María and Tellería 2015). More research on juveniles and small mammals is needed (Table 2). We are also at the stage of considering interventions to improve the welfare of some wild mammals and birds, ranging from pigeon fertility control to reducing feral cat predation. To make these interventions as effective as possible, it would be valuable to understand the replaceability of different sources of mortality.

Table 2
Research gaps and proposed priorities by taxon.
Fish and juvenile amphibians are still seriously lacking in data on cause-specific mortality, especially relating to juveniles. Research on both of these groups is challenged by the fact that they spend some or all of their lives underwater, but biologging technologies — including pop-up satellite archival tags (PSATs) (Tolentino et al. 2017) and acoustic telemetry (Weinz et al. 2020) — could enable more research on cause of death in aquatic environments, especially related to predation.
A surprising amount is known about cause-specific mortality in insects regarded as agricultural pests (Table 1). However, very little research has been done on the deaths of insects with less agricultural relevance. From the existing research, it is clear that insects face challenges that would surprise us. For example, Asiimwe et al. (2006) found that many whiteflies died as a result of poorly understood developmental abnormalities, such as failure to complete metamorphosis. Drowning during rainy weather was one of the foremost causes of death among the moths of Pereira et al. (2007), and plant chemical defenses were responsible for a majority of the deaths of wheat stem sawflies monitored by Acchami et al. (2020).
As many challenges as we know that animals face in the wild, some may yet be masked by the urgent threats they face from humans through hunting, fishing, extermination, and competition for resources. Humans should leave more space for wildlife, and try not to make their existence harder than it already is, but we should also be willing to go further. Learning more about cause-specific mortality in different populations and environmental contexts can inform plans for the most effective actions to improve wild animal welfare.
References
Achhami, Buddhi B., Robert K. D. Peterson, Jamie D. Sherman, Gadi V. P. Reddy, and David K. Weaver. 2020. ‘Multiple Decrement Life Tables of Cephus Cinctus Norton (Hymenoptera: Cephidae) across a Set of Barley Cultivars: The Importance of Plant Defense versus Cannibalism’. PLOS ONE 15 (9): e0238527. https://doi.org/10.1371/journal.pone.0238527.
Andis, M. D., and C. L. Meek. 1985. ‘Mortality and Survival Patterns for the Immature Stages of Psorophora Columbiae’. Journal of the American Mosquito Control Association 1 (3): 357–62.
Asiimwe, P., J. S. Ecaat, M. Otim, D. Gerling, S. Kyamanywa, and J. P. Legg. 2007. ‘Life-Table Analysis of Mortality Factors Affecting Populations of Bemisia Tabaci on Cassava in Uganda’. Entomologia Experimentalis et Applicata 122 (1): 37–44. https://doi.org/10.1111/j.1570-7458.2006.00487.x.
Bartmann, Richard M., Gary C. White, and Len H. Carpenter. 1992. ‘Compensatory Mortality in a Colorado Mule Deer Population’. Wildlife Monographs, no. 121: 3–39.
Benelli, G., Pozzebon, A., & Reaz, M. B. I. 2013. ‘RFID under water: Technical issues and applications’ (pp. 379-395). Rijeka: InTech. https://doi.org/10.5772/53934.
Bergman, Eric J., Paul F. Doherty, Gary C. White, and A. Andrew Holland. 2015. ‘Density Dependence in Mule Deer: A Review of Evidence’. Wildlife Biology 21 (1): 18–29. https://doi.org/10.2981/wlb.00012.
Buenestado, F. J., Ferreras, P., BLANCO‐AGUIAR, J. A., Tortosa, F. S., & Villafuerte, R. 2009. ‘Survival and causes of mortality among wild Red‐legged Partridges Alectoris rufa in southern Spain: implications for conservation’. Ibis 151 (4): 720-730. https://doi.org/10.1111/j.1474-919X.2009.00952.x.
Butler, J. A., & Sowell, S. 1996. ‘Survivorship and predation of hatchling and yearling gopher tortoises, Gopherus polyphemus’. Journal of Herpetology 30 (3): 455-458. https://doi.org/10.2307/1565195.
Chomel, B.B., R.W. Kasten, G. Chappuis, M. Soulier, and Y. Kikuchi. 1998. ‘Serological Survey of Selected Canine Viral Pathogens and Zoonoses in Grizzly Bears (Ursus Arctos Horribilis) and Black Bears (Ursus Americanus) from Alaska: -EN- -FR- -ES-’. Revue Scientifique et Technique de l’OIE 17 (3): 756–66. https://doi.org/10.20506/rst.17.3.1134.
Collins, C., and R. Kays. 2011. ‘Causes of Mortality in North American Populations of Large and Medium-Sized Mammals’. Animal Conservation 14 (5): 474–83. https://doi.org/10.1111/j.1469-1795.2011.00458.x.
Cooley, Hilary S., Robert B. Wielgus, Gary M. Koehler, Hugh S. Robinson, and Benjamin T. Maletzke. 2009. ‘Does Hunting Regulate Cougar Populations? A Test of the Compensatory Mortality Hypothesis’. Ecology 90 (10): 2913–21. https://doi.org/10.1890/08-1805.1.
Crosby, Lyle A., and Randy Graham. 1986. ‘Population Dynamics and Expansion Rates of Black-Tailed Prairie Dogs’. Proceedings of the Vertebrate Pest Conference 12 (12). https://escholarship.org/uc/item/23t218h1.
Dieperink, C., S. Pedersen, and M. I. Pedersen. 2001. ‘Estuarine Predation on Radiotagged Wild and Domesticated Sea Trout (Salmo Trutta L.) Smolts’. Ecology of Freshwater Fish 10 (3): 177–83. https://doi.org/10.1034/j.1600-0633.2001.100307.x.
Fritts, Anthony L., and Todd N. Pearsons. 2004. ‘Smallmouth Bass Predation on Hatchery and Wild Salmonids in the Yakima River, Washington’. Transactions of the American Fisheries Society 133 (4): 880–95. https://doi.org/10.1577/T03-003.1.
Hill, Jacob E., Travis L. DeVault, and Jerrold L. Belant. 2019. ‘Cause-Specific Mortality of the World’s Terrestrial Vertebrates’. Global Ecology and Biogeography 28 (5): 680–89. https://doi.org/10.1111/geb.12881.
Himes, J. G., Hardy, L. M., Rudolph, D. C., & Burgdorf, S. J. 2002. ‘Growth rates and mortality of the Louisiana pine snake (Pituophis ruthveni)’. Journal of Herpetology 36 (4): 683-687. https://doi.org/10.2307/1565941.
Hostetter, Nathan J., Allen F. Evans, Daniel D. Roby, and Ken Collis. 2012. ‘Susceptibility of Juvenile Steelhead to Avian Predation: The Influence of Individual Fish Characteristics and River Conditions’. Transactions of the American Fisheries Society 141 (6): 1586–99. https://doi.org/10.1080/00028487.2012.716011.
Hurst, T. P. 2007. ‘Causes and Consequences of Winter Mortality in Fishes’. Journal of Fish Biology 71 (2): 315–45. https://doi.org/10.1111/j.1095-8649.2007.01596.x.
Jones, Davy. 1982. ‘Predators and Parasites of Temporary Row Crop Pests: Agents of Irreplaceable Mortality or Scavengers Acting Prior to Other Mortality Factors?’ Entomophaga 27 (3): 245–65. https://doi.org/10.1007/BF02374809.
Karut, K., and S. E. Naranjo. 2009. ‘Mortality Factors Affecting Bemisia Tabaci Populations on Cotton in Turkey’. Journal of Applied Entomology 133 (5): 367–74. https://doi.org/10.1111/j.1439-0418.2008.01369.x.
Kays, Roland, Margaret C. Crofoot, Walter Jetz, and Martin Wikelski. 2015. ‘Terrestrial Animal Tracking as an Eye on Life and Planet’. Science 348 (6240). https://doi.org/10.1126/science.aaa2478.
Klaassen, R. H., Hake, M., Strandberg, R., Koks, B. J., Trierweiler, C., Exo, K. M., ... & Alerstam, T. 2014. ‘When and where does mortality occur in migratory birds? Direct evidence from long‐term satellite tracking of raptors’. Journal of Animal Ecology 83 (1): 176-184. https://doi.org/10.1111/1365-2656.12135.
Koed, Anders, Henrik Baktoft, and Brian Daniel Bak. 2006. ‘Causes of Mortality of Atlantic Salmon (Salmo Salar) and Brown Trout (Salmo Trutta) Smolts in a Restored River and Its Estuary’. River Research and Applications 22 (1): 69–78. https://doi.org/10.1002/rra.894.
Krkošek, Martin. 2017. ‘Population Biology of Infectious Diseases Shared by Wild and Farmed Fish1’. Canadian Journal of Fisheries and Aquatic Sciences, January. https://doi.org/10.1139/cjfas-2016-0379.
Krumm, Caroline E., Mary M. Conner, N. Thompson Hobbs, Don O. Hunter, and Michael W. Miller. 2010. ‘Mountain Lions Prey Selectively on Prion-Infected Mule Deer’. Biology Letters 6 (2): 209–11. https://doi.org/10.1098/rsbl.2009.0742.
LeCount, Albert L. 1987. ‘Causes of Black Bear Cub Mortality’. Bears: Their Biology and Management 7: 75–82. https://doi.org/10.2307/3872610.
Lenting, B., B. Gartrell, A. Kokosinska, P. J. Duignan, S. Michael, S. Hunter, and W. D. Roe. 2019. ‘Causes of Adult Mortality in Two Populations of New Zealand Sea Lions (Phocarctos Hookeri)’. Veterinary and Animal Science 7 (June): 100057. https://doi.org/10.1016/j.vas.2019.100057.
Naranjo, Steven E., and Peter C. Ellsworth. 2005. ‘Mortality Dynamics and Population Regulation in Bemisia Tabaci’. Entomologia Experimentalis et Applicata 116 (2): 93–108. https://doi.org/10.1111/j.1570-7458.2005.00297.x.
Palma, L., Beja, P., Pais, M., & Cancela Da Fonseca, L. 2006. ‘Why do raptors take domestic prey? The case of Bonelli's eagles and pigeons’. Journal of Applied Ecology 43 (6): 1075-1086. https://doi.org/10.1111/j.1365-2664.2006.01213.x.
Pereira, E. J. G., M. C. Picanço, L. Bacci, A. L. B. Crespo, and R. N. C. Guedes. 2007. ‘Seasonal Mortality Factors of the Coffee Leafminer, Leucoptera Coffeella’. Bulletin of Entomological Research 97 (4): 421–32. https://doi.org/10.1017/S0007485307005202.
Peterson, Robert K. D., Ryan S. Davis, Leon G. Higley, and Odair A. Fernandes. 2009. ‘Mortality Risk in Insects’. Environmental Entomology 38 (1): 2–10. https://doi.org/10.1603/022.038.0102.
Raby, Graham D., Jessica R. Packer, Andy J. Danylchuk, and Steven J. Cooke. 2014. ‘The Understudied and Underappreciated Role of Predation in the Mortality of Fish Released from Fishing Gears’. Fish and Fisheries 15 (3): 489–505. https://doi.org/10.1111/faf.12033.
Roux, Olivier, and Johann Baumgärtner. 1998. ‘Evaluation of Mortality Factors and Risk Analysis for the Design of an Integrated Pest Management System’. Ecological Modelling 109 (1): 61–75. https://doi.org/10.1016/S0304-3800(98)00035-0.
Sáenz-de-Santa-María, A., & Tellería, J. L. 2015. ‘Wildlife-vehicle collisions in Spain’. European Journal of Wildlife Research 61 (3): 399-406. https://doi.org/10.1007/s10344-015-0907-7.
Siler, W. 1979. ‘A competing‐risk model for animal mortality’. Ecology 60 (4): 750-757. https://doi.org/10.2307/1936612.
Stapp, Paul, Michael F. Antolin, and Mark Ball. 2004. ‘Patterns of Extinction in Prairie Dog Metapopulations: Plague Outbreaks Follow El Ninì Events’. Frontiers in Ecology and the Environment 2 (5): 235–40. https://doi.org/10.1890/1540-9295(2004)002[0235:POEIPD]2.0.CO;2.
Stenkat, J., Krautwald-Junghanns, M. E., & Schmidt, V. 2013. ‘Causes of morbidity and mortality in free-living birds in an urban environment in Germany’. Ecohealth 10 (4): 352-365. https://doi.org/10.1007/s10393-013-0868-9.
Sterud, E, T Forseth, O Ugedal, Tt Poppe, A Jørgensen, T Bruheim, H Fjeldstad, and Ta Mo. 2007. ‘Severe Mortality in Wild Atlantic Salmon Salmo Salar Due to Proliferative Kidney Disease (PKD) Caused by Tetracapsuloides Bryosalmonae (Myxozoa)’. Diseases of Aquatic Organisms 77 (October): 191–98. https://doi.org/10.3354/dao01846.
Tolentino, Emily R., Russell P. Howey, Lucy A. Howey, Lance K. B. Jordan, R. Dean Grubbs, Annabelle Brooks, Sean Williams, Edward J. Brooks, and Oliver N. Shipley. 2017. ‘Was My Science Project Eaten? A Novel Approach to Validate Consumption of Marine Biologging Instruments’. Animal Biotelemetry 5 (1): 3. https://doi.org/10.1186/s40317-016-0117-4.
Weinz, Amy A., Jordan K. Matley, Natalie V. Klinard, Aaron T. Fisk, and Scott F. Colborne. 2020a. ‘Identification of Predation Events in Wild Fish Using Novel Acoustic Transmitters’. Animal Biotelemetry 8 (1): 28. https://doi.org/10.1186/s40317-020-00215-x.
Methods for studying wild animals’ causes of death
In part two of this series, Luke Hecht introduces some of the tools and approaches used to study wild animals’ causes of death.
Key takeaways
- Cause of death is an important area of wild animal welfare research.
- Accurate cause-specific mortality rates are difficult to obtain.
- Multistate modeling can provide insight where information about cause-specific mortality rates is incomplete.
- Experimental studies and improved monitoring technology can improve our ability to diagnose cause of death, estimate cause-specific mortality rates, and understand compensatory mortality.
- Future research should identify and focus on the causes of death that involve the most suffering.
In a world with perfect monitoring of wild populations, biologists could be immediately alerted to the death of an animal, show up on the scene, and perform a necropsy to determine the animal’s precise cause of death. They would also know everything about the age and health history of this animal, having monitored them from birth. Unfortunately, it is not practical to monitor most wild populations at nearly this level of detail. Our understanding of how and why wild animals die must be pieced together from statistical models and sparse empirical data. In this post, I will introduce some of the tools and approaches used to study wild animals’ causes of death.
Death assemblages
The most straightforward way to study wild animals’ causes of death is to scour a habitat for corpses. The resulting collection is sometimes called a “death assemblage.” When the body of a recently deceased animal is recovered, indicators such as injuries, signs of disease, and stomach contents may make it possible to determine what killed them (Behrensmeyer and Miller 2012).
Determining cause of death from serendipitously recovered corpses depends on finding animals soon after their deaths. Death assemblage surveys are therefore most useful for animals whose remains are more likely to be discovered, like coastal marine mammals. Their large size means they take a while to decay, and their bodies often wash up on beaches where they are highly visible and quickly reported to authorities or researchers (Tinker et al. 2016). Recovery of well-preserved corpses is much less common for animals who are small or live in lush environments like forests, which typically provide more concealment and support more rapid decomposition.
The probability of recovering a corpse can also be biased by whatever caused the animal’s death (Gerber et al. 2004). For example, the bodies of animals who were killed by predators may be less likely to be recovered because they are almost entirely consumed on the spot or are carried away from the site (e.g. hyenas stashing bones of prey, Kuhn 2005). On the other hand, animals who die from starvation, disease, old age, or something else that leads to their gradual weakening may attempt to conceal themselves in their final hours. While their bodies are likely to remain intact for longer, they may go unnoticed by researchers.
Certain causes of death are also inherently more difficult to diagnose, so researchers need to be wary of simply ignoring data in an “unknown” category that may be disproportionately filled with instances of hard-to-distinguish causes of death. For example, in a survey of bones found in a hyena den, presumably belonging to their past prey, Kuhn (2005) identified roughly a quarter of the specimens to species level. Without further information, any quantitative claims about the composition of the local ecosystem based on these data would have to be taken with serious caution because the remaining 75% of “unknown” specimens might be drawn unequally from underlying populations. For example, animals who are morphologically similar to other species, or animals who are not preferred prey for hyenas, might be underrepresented in resulting estimates.
Bias is a pervasive problem in cause of death surveys. A naive survey of rabbit corpses in a particular forest might conclude that most recent deaths were attributable to some rapidly debilitating disease, while in fact the majority have been carried off by birds of prey without a trace. To estimate the true rates of alternative causes of death from corpse recovery data, researchers need to account for the fact that the forensic clues to certain causes of death may be more or less detectable than others, depending on a host of circumstances.
Accounting for incomplete information
The inevitable incompleteness of corpse recovery surveys can be partially offset by explicitly modeling sources of uncertainty along the chain of events from death by a specific cause, to corpse recovery, to cause identification. This approach is sometimes called multistate modeling (Hougaard 1999; Gauthier and Lebreton 2009). These models make a distinction between the objective chain of events that occurred to an individual animal and the observable evidence of those events (Koons et al. 2014). For example, researchers generally do not directly observe rodents being preyed on by raptors. A fraction of predation events leave behind bits of fur or the partially eaten body of the rodent. Researchers then have a particular probability of encountering these bits of evidence and deducing the correct cause of death.
Box 1
To illustrate what a multistate model looks like in practice, imagine that a flock of birds begins to roost every day in a tree outside where you live. The foliage is dense, so you cannot see all of the roosting birds at any one time through your window. One of the birds (“Bird X”) has distinctive markings, with which you can usually distinguish them from the others. One day, you can’t see Bird X in the tree. Your first worry is that Bird X might have died, but then you remember that they might just be out of view behind a branch. A week passes and Bird X still hasn’t been seen in the tree. Is it more likely that you simply have not caught sight of Bird X for seven days in a row, or that Bird X died sometime in the week?
This problem can be solved by estimating the exact proportion of birds in the tree who can be seen on any given day (“detection probability”), the probability of recognizing the identity of Bird X if you set eyes on them (“identification probability”), and the average daily survival probability for each bird in this population. These can then be incorporated into a model where Bird X is considered to be in one of two states: alive or dead. Using the model, we can determine whether our series of observations of the tree outside is better explained by Bird X being alive or by being dead. Marescot et al. (2015) applied a similar sort of model to observations of tagged black-tailed deer to estimate their age-specific survival rates and the impact of predation on their population growth.
While multistate modeling is frequently used to estimate survival rates in wild animal populations, similarly nuanced studies of cause of death are rare. This may reflect both a general disinterest in cause-of-death research and a greater confidence in the raw data that does exist. Studying human-caused deaths or species for which there is a good infrastructure for monitoring removes some uncertainty and lessens the need for multistate models. These statistical methods are most important to apply when studying populations where the vast majority of individuals’ deaths go undocumented, leaving room for large biases in the data. For example, Gerber et al. (2004) investigated cause-specific mortality of sea otters. To do so, they took advantage of an extensive network of marine mammal research groups to recover and analyze carcasses that washed up on the beach. Using a demographic model, they estimated that their sample represented just under half of all mortality during the study period. To sample half of all deceased individuals in a wild population of marine mammals is a remarkable achievement, but it highlights how much potential there is for unequal cause-specific recovery rates to produce biased results, especially in less intensively monitored populations. In marine mammals, for example, different predator species may hunt at different distances from shore, or near rocky cliffs as opposed to smooth beaches. Either of those factors could affect the probability of a carcass washing up onshore and being recovered by researchers (Joly et al. 2009).
The biggest weakness of purely observational studies of wild animal mortality is that it is difficult to know what data are missing. The simplest way of overcoming this is to tag a set number of animals at the beginning of a study and only count the remains of tagged individuals. This sort of study design is still subject to biased recovery rates, but the magnitude of bias is limited because corpses of untagged individuals are excluded. Schaub and Pradel (2004) used such an approach to estimate the proportion of white storks killed in airborne collisions with power lines in Switzerland (Figure 1a). They found that simply counting corpses overestimated the proportion of juvenile white storks killed by power lines by about 25%. It makes sense that the bodies of birds killed by power line collisions would be more conspicuous than birds who die otherwise, because the ground around power lines tends to be more exposed to allow for maintenance access. Power lines may also serve as natural concentrating points for associated deaths, whereas deaths by other causes may be uniformly distributed across the landscape.
Limiting analyses to tagged animals is effective at preventing overestimation of particular causes of death, but it can also leave researchers with a lot of missing data from individuals whose bodies were not recovered or who lost their tags. Steps can be taken to improve the recovery rate of tagged animals, such as attaching radio trackers to animals alongside conventional tags (Tavecchia et al. 2011; Figure 1b). Records of when a tagged animal was last seen alive can also complement corpse recovery data, because they allow for independent estimation of survival rates and rates of tag loss. Without this information, corpse recovery data alone cannot distinguish between an animal who is still alive and one who died but was never recovered. Fortunately, technical improvements in the precision and durability of radio trackers are making it practical to do this remotely. Even with relatively basic GPS tags, statistical models can distinguish tag loss or technical failure from actual death of the tagged animal with almost perfect accuracy (Sergio et al. 2018). Electronic tags capable of recording animals’ fine-scale movements or vital signs can also provide evidence related to an animal’s cause of death, such as how sudden it was or what posture the animal adopted in their final moments (Cooke 2008; Brown et al. 2013). Advances in the field of animal remote sensing will be helpful for learning about cause-specific mortality at a level of detail that is currently impossible for researchers to observe.

Figure 1
Multistate models treat wild animals’ fates — observed or unobserved — as the product of a series of conditional probabilities reflecting the combination of events over their life. A) A basic model that considers whether an animal is alive or dead, what caused their death, and whether their corpse was recovered (Schaud and Pradel 2004). B) A more complex model that estimates the probabilities of a bird dying from poisoning or electrocution, and ultimately the probability of a given bird being sighted again alive or dead (Tavecchia et al. 2011). In both of these models, the state transition probabilities are estimated based on the history of encounters between researchers and each tagged animal over the course of a study. For example, if researchers observe an individual after not sighting them for multiple field seasons, then that sighting might both increase the estimated survival rate and decrease the estimated detection rate.
Experimental study of cause-specific mortality
Rather than simply observing the fates of wild animals, experimental approaches directly exclude or introduce specific causes of death. The most common form of this approach is to exclude predators with fencing or other deterrents. For example, Smith et al. (2012) excluded all medium-sized mammalian predators from some nesting grounds of the gopher tortoise, but not others, and found that tortoises born in the predator exclusion zones were almost twice as likely to still be alive one year later. Similarly, Conner et al. (2011) found that exclusion fencing reduced the number of cotton rats killed by mammals. However, increased predation by birds and snakes largely made up for the decreased predation by mammals, especially after a forest fire destroyed much of the undergrowth that could otherwise conceal the rats.
Although most experimental research on cause-specific mortality to date has focused on hunting or predation, a similar approach could be used to study the effectiveness of interventions against other common causes of death, including curable diseases or hunger (c.f. Rickett et al. 2013). However, interventions like supplemental feeding, disease prevention and predator exclusion may result in compensatory mortality, where individuals spared from one cause of death are at elevated risk of dying soon after because of another cause. The most effective way of learning about compensatory mortality risk is to use an experimental rather than observational study design, in conjunction with the same best practices for observational studies as described previously (Bergman et al. 2015).
Outlook for welfare-focused cause of death research
From the perspective of wild animal welfare, understanding cause-specific mortality is morally urgent. A few shifts in the current approach to cause-of-death research will help us determine which issues to prioritize to help animals. Most importantly, much more research should be focused on the most common deaths in the wild: deaths of juvenile animals and members of species with large global populations. Because they are so numerous, these individuals’ experiences of dying are especially important to understand if we value all sentient animals’ welfare equally. These animals tend to be more difficult to monitor due to their smaller body size, but studying their mortality is becoming ever easier with new technology. At the current rate of progress, organisms weighing around one gram could be tracked via GPS within a decade or less (Kays et al. 2015).
One major source of uncertainty in wild animal mortality research is the diagnosis of an animal’s cause of death. This is especially true for low-tech studies, where corpses are encountered by chance, rather than sought at a particular location based on radio tracking and a mortality signal. As a result, corpses may be significantly decomposed or scavenged. Even under ideal circumstances, it can be difficult to determine the order in which apparent injuries occurred. In lieu of perfect monitoring, researchers could perform detailed analysis on a subset of the recovered bodies to evaluate the reliability of their field diagnoses of the rest, as Joly et al. (2009) did with sea otter carcasses. Studies could also be dedicated to intensive monitoring of a relatively small number of animals so that when they die the cause can be known with high confidence and researchers can quickly find their corpses to document field-identifiable characteristics and how these change over the coming days or weeks, as well as estimate recovery rates. The field of taphonomy has long filled a similar supportive role for paleobiology (Behrensmeyer and Miller 2012). Upfront investment in this sort of meta-research could enable subsequent studies of wild animal mortality to accomplish more with less (c.f. Loannidis 2018).
Welfare-focused researchers should aim to identify and focus on the causes of death that entail much more suffering than others. This could involve a reframing of cause-specific mortality research where rather than asking how animals of a given species are likely to die, researchers ask who the victims of a particular cause of death are likely to be. Rather than tracking marmots and determining what proportion were preyed on by cougars, for instance, researchers could monitor cougars using cameras or fecal analysis to learn who their prey were (Popanom et al. 2011). This sort of cause-centric approach might also be appropriate in contexts where a single ultimate cause can lead to several different proximate causes of death which vary in their severity. For example, researchers could survey a forest in the aftermath of a fire and try to determine what proportion of fatalities occurred by burning, asphyxiation, subsequent starvation, or other proximate causes related to the fire (Gutiérrez and Javier de Miguel, 2020). Simply verifying that a certain cause of death occurred at least once in a given population may be useful to inform future work. If so, low-cost cause-specific mortality studies could use a form of rarefaction analysis to judge how comprehensively the causes of death present in a population have been sampled, with a focus on enumerating the causes rather than estimating their relative frequencies.
Efforts to improve the health and lifespans of our fellow humans are supported by publicly available data on the most common causes of death. This is part of how governments decide which public health interventions and fields of medical research to allocate funding to. A comparable wealth of data could help us identify which wild animals need the most urgent aid, and what form that aid should take. In the next part of this series, I will give an overview of some of the data that does exist on the causes of wild animal mortality.
References
Behrensmeyer, Anna K., and Joshua H. Miller. 2012. ‘Building Links Between Ecology and Paleontology Using Taphonomic Studies of Recent Vertebrate Communities’. In Paleontology in Ecology and Conservation, edited by Julien Louys, 69–91. Springer Earth System Sciences. Berlin, Heidelberg: Springer. https://doi.org/10.1007/978-3-642-25038-5_5.
Bergman, Eric J., Paul F. Doherty, Gary C. White, and A. Andrew Holland. 2015. ‘Density Dependence in Mule Deer: A Review of Evidence’. Wildlife Biology 21 (1): 18–29. https://doi.org/10.2981/wlb.00012.
Brown, Danielle D., Roland Kays, Martin Wikelski, Rory Wilson, and A. Peter Klimley. 2013. ‘Observing the Unwatchable through Acceleration Logging of Animal Behavior’. Animal Biotelemetry 1 (1): 20. https://doi.org/10.1186/2050-3385-1-20.
Conner, L. Mike, Steven B. Castleberry, and Anna M. Derrick. 2011. ‘Effects of Mesopredators and Prescribed Fire on Hispid Cotton Rat Survival and Cause-Specific Mortality’. The Journal of Wildlife Management 75 (4): 938–44. https://doi.org/10.1002/jwmg.110.
Cooke, Steven J. 2008. ‘Biotelemetry and Biologging in Endangered Species Research and Animal Conservation: Relevance to Regional, National, and IUCN Red List Threat Assessments’. Endangered Species Research 4 (1–2): 165–85. https://doi.org/10.3354/esr00063.
Gauthier, Gilles, and Jean-Dominique Lebreton. 2008. ‘Analysis of Band-Recovery Data in a Multistate Capture-Recapture Framework’. Canadian Journal of Statistics 36 (1): 59–73. https://doi.org/10.1002/cjs.5550360107.
Gerber, Leah R., M. Tim Tinker, Daniel F. Doak, James A. Estes, and David A. Jessup. 2004. ‘Mortality Sensitivity in Life-Stage Simulation Analysis: A Case Study of Southern Sea Otters’. Ecological Applications 14 (5): 1554–65. https://doi.org/10.1890/03-5006.
Gutiérrez, J. and de Miguel, F. J. 2020. Challenges posed by fires to wild animals and how to help: A literature review, Oakland: Animal Ethics, retrieved from https://www.animalethics.org/fire-wild-animals-help.
Hougaard, Philip. 1999. ‘Multi-State Models: A Review’. Lifetime Data Analysis 5 (3): 239–64. https://doi.org/10.1023/A:1009672031531.
Ioannidis, John P. A. 2018. ‘Meta-Research: Why Research on Research Matters’. PLOS Biology 16 (3): e2005468. https://doi.org/10.1371/journal.pbio.2005468.
Joly, Damien O., Dennis M. Heisey, Michael D. Samuel, Christine A. Ribic, Nancy J. Thomas, Scott D. Wright, and Irene E. Wright. 2009. ‘Estimating Cause-Specific Mortality Rates Using Recovered Carcasses’. Journal of Wildlife Diseases 45 (1): 122–27. https://doi.org/10.7589/0090-3558-45.1.122.
Kays, R., M. C. Crofoot, W. Jetz, and M. Wikelski. 2015. ‘Terrestrial Animal Tracking as an Eye on Life and Planet’. Science 348 (6240): aaa2478–aaa2478. https://doi.org/10.1126/science.aaa2478.
Koons, David N., Marlène Gamelon, Jean-Michel Gaillard, Lise M. Aubry, Robert F. Rockwell, François Klein, Rémi Choquet, and Olivier Gimenez. 2014. ‘Methods for Studying Cause-Specific Senescence in the Wild’. Methods in Ecology and Evolution 5 (9): 924–33. https://doi.org/10.1111/2041-210X.12239.
Kuhn, Brian. 2005. ‘The Faunal Assemblages and Taphonomic Signatures of Five Striped Hyaena (Hyaena Hyaena Syriaca) Dens in the Desert of Eastern Jordan’. Levant 37 (1): 221–34. https://doi.org/10.1179/lev.2005.37.1.221.
Marescot, Lucile, Tavis D. Forrester, David S. Casady, and Heiko U. Wittmer. 2015. ‘Using Multistate Capture–Mark–Recapture Models to Quantify Effects of Predation on Age-Specific Survival and Population Growth in Black-Tailed Deer’. Population Ecology 57 (1): 185–97. https://doi.org/10.1007/s10144-014-0456-z.
Pompanon, Francois, Bruce E. Deagle, William O. C. Symondson, David S. Brown, Simon N. Jarman, and Pierre Taberlet. 2012. ‘Who Is Eating What: Diet Assessment Using Next Generation Sequencing’. Molecular Ecology 21 (8): 1931–50. https://doi.org/10.1111/j.1365-294X.2011.05403.x.
Rickett, Jennifer, Cody J. Dey, Jillian Stothart, Constance M. O’Connor, James S. Quinn, and Weihong Ji. 2013. ‘The Influence of Supplemental Feeding on Survival, Dispersal and Competition in Translocated Brown Teal, or Pateke (Anas Chlorotis)’. Emu - Austral Ornithology 113 (1): 62–68. https://doi.org/10.1071/MU12053.
Schaub, Michael, and Roger Pradel. 2004. ‘Assessing the Relative Importance of Different Sources of Mortality from Recoveries of Marked Animals’. Ecology 85 (4): 930–38. https://doi.org/10.1890/03-0012.
Sergio, Fabrizio, Alessandro Tanferna, Julio Blas, Guillermo Blanco, and Fernando Hiraldo. 2019. ‘Reliable Methods for Identifying Animal Deaths in GPS- and Satellite-Tracking Data: Review, Testing, and Calibration’. Journal of Applied Ecology 56 (3): 562–72. https://doi.org/10.1111/1365-2664.13294.
Smith, Lora L., David A. Steen, L. Mike Conner, and Jessica C. Rutledge. 2013. ‘Effects of Predator Exclusion on Nest and Hatchling Survival in the Gopher Tortoise’. The Journal of Wildlife Management 77 (2): 352–58. https://doi.org/10.1002/jwmg.449.
Tavecchia, Giacomo, Jaume Adrover, Antoni Muñoz Navarro, and Roger Pradel. 2012. ‘Modelling Mortality Causes in Longitudinal Data in the Presence of Tag Loss: Application to Raptor Poisoning and Electrocution’. Journal of Applied Ecology 49 (1): 297–305. https://doi.org/10.1111/j.1365-2664.2011.02074.x.
Tinker, M. Tim, Brian B. Hatfield, Michael D. Harris, and Jack A. Ames. 2016. ‘Dramatic Increase in Sea Otter Mortality from White Sharks in California’. Marine Mammal Science 32 (1): 309–26. https://doi.org/10.1111/mms.12261.
Why cause of death matters for wild animal welfare
In part one of this series, Luke Hecht introduces approaches to studying wild animals’ causes of death, with the goal of making work in this field maximally useful for understanding wild animal welfare.
The most fundamental reason for viewing death as a bad thing is that it deprives individuals of future experiences, assuming those experiences would have been predominantly positive. However, the process of dying is also often painful in itself, and an individual’s death can have additional negative impacts—emotional and material—on others, including family and the broader population.
The costs of a death depend on its cause. Humans are willing to sacrifice periods of healthy life to avoid especially unpleasant deaths, and see various causes of death as more fearsome than others (Sunstein 1997; Chapple et al. 2006). Of course, people’s perception of how much suffering particular deaths would entail may be biased by things like squeamishness; they may assume that because a death is gruesome it must be especially painful. The terminal stages in the process of dying, during which suffering presumably outweighs pleasure, comprise a small fraction of most humans’ lives. Willingness to trade years of healthy life to obtain a quicker, less painful death implies that either the original manner of death must be extremely bad, or that we have an exaggerated sense of how bad dying is.
As recently as the 19th century, only around half of newborns could expect to survive to adulthood (Riley 2005). Many human cultures have had traditions of only conferring names on children when they reached a certain age, potentially because it was understood that most would not survive that long (but that if they did, they would have a decent prospect of reaching adulthood) (Lancy 2014). For example, birth registration was not a legal requirement in the United Kingdom until 1874 (ONS 2015). As average lifespan increases, the dying process represents an ever smaller proportion of human life.
While human childhood mortality has declined dramatically throughout much of the world, most wild animals still die at a young age relative to the longest-lived members of their own species (Figure 1). For these animals, the process of dying may actually represent a substantial portion of their lifetimes, suggesting that, in addition to knowing the most likely causes of death for entire populations, it would be especially valuable to know which manners of death are common among juveniles in specific wild animal populations.
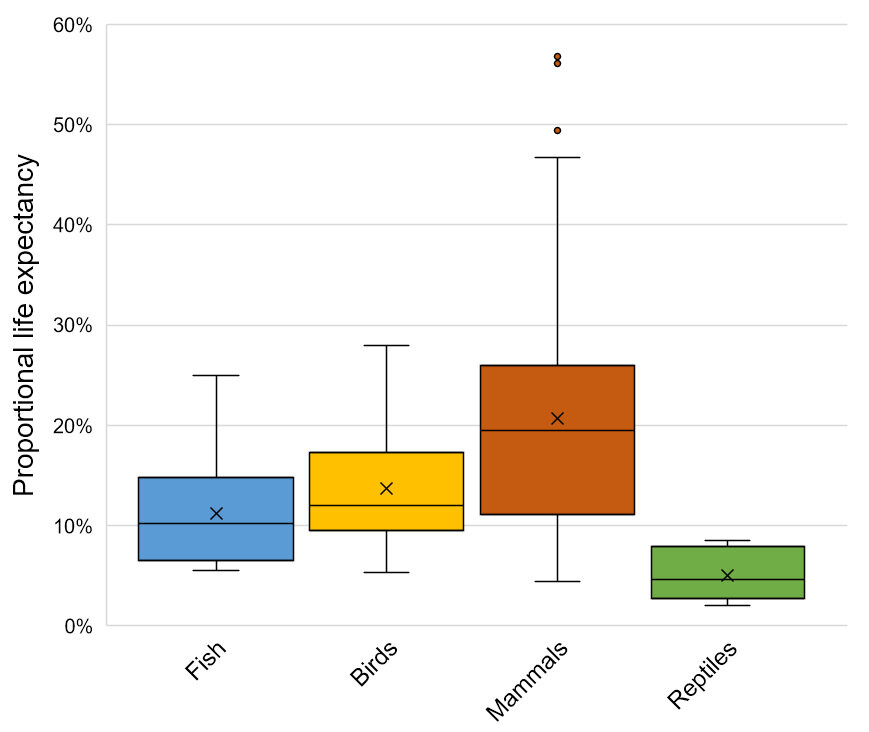
Figure 1: A boxplot showing life expectancy as a percentage of a species’ maximum lifespan for 152 populations of fish (n=16), birds (n=54), mammals (n=72) and reptiles (n=10). Life expectancies were calculated from models found in the COMADRE database (Salguero-Gomez et al. 2016), and maximum lifespans were obtained from AnAge (De Magalhães et al. 2005). Across major vertebrate classes, most individuals live to only 10-30% of the age of the oldest known individuals of their species.
“Cause” of death can be interpreted in at least two different ways: “manner” of death, or “ultimate cause” of death. Both matter for wild animal welfare, but they can be difficult to disentangle in practice. For example, an animal who died during a forest fire may have died by burning or asphyxiation. Other animals may die in the immediate aftermath of a forest fire from dehydration or infected burn wounds. The welfare impact of these different manners of death would likely vary, and it is possible in theory to rank their welfare impacts based on the intensity and duration of suffering leading up to death (Figure 2; Sharp and Saunders 2011). Learning about the precise manner in which wild animals die can help us prioritize hazards to protect them from to reduce instances of the most extreme suffering. On the other hand, it is often easier to estimate the number of animals who died due to an ultimate cause - such as forest fires - than to break this down into specific manners of death. Learning about an ultimate cause of death, and the number of deaths resulting from it, is also useful for prioritizing welfare interventions directed at extending good lives.

Figure 2: An example scheme for grading the welfare impact of different manners of death, taking into account both the severity of suffering inflicted and the length of time it takes for the animal to die. Adapted from work by Sharp and Saunders (2011) on wild animal culling methods.
To minimize the suffering animals experience in the wild, we need to understand how and why wild animals die, paying particular attention to the most numerous experiences: that is, the deaths of juvenile animals belonging to common species. The rates of different causes of death are also valuable to know because even if dying turns out to be a minor contributor to lifetime welfare, or the difference in severity between manners of death is relatively low, substituting one manner of death for another less painful one could improve individual welfare without affecting population size, minimizing the number of variables we need to consider to determine whether an intervention is worth implementing.
In a series of posts to follow, I will review the methods available for studying causes of wild animal deaths and present a selection of published data on cause-specific mortality rates. Finally, I’ll describe some original modeling by former Wild Animal Initiative Intern Anthony DiGiovanni that explicitly considers how cause of death may change along with the size and demography of wild animal populations.
References
Sunstein, C. (1997). Bad deaths. Journal of Risk and Uncertainty, 14(3), 259-282.
Chapple, A., Ziebland, S., McPherson, A., & Herxheimer, A. (2006). What people close to death say about euthanasia and assisted suicide: a qualitative study. Journal of Medical Ethics, 32(12), 706-710.
Riley, J. C. (2005). Estimates of regional and global life expectancy, 1800–2001. Population and Development Review, 31(3), 537-543.
Lancy, D. (2014). “Babies aren’t persons”: A survey of delayed personhood. In H. Otto & H. Keller (Eds.), Different Faces of Attachment: Cultural Variations on a Universal Human Need (pp. 66-110). Cambridge: Cambridge University Press.
Office for National Statistics. (2015). How has life expectancy changed over time? Retrieved from https://www.ons.gov.uk/peoplepopulationandcommunity/birthsdeathsandmarriages/lifeexpectancies/articles/howhaslifeexpectancychangedovertime/2015-09-09.
Sharp, T., & Saunders, G. (2011). A model for assessing the relative humaneness of pest animal control methods. Canberra, Australia: Department of Agriculture, Fisheries and Forestry.
De Magalhaes, J. P., & Costa, J. (2009). A database of vertebrate longevity records and their relation to other life‐history traits. Journal of Evolutionary Biology, 22(8), 1770-1774.
Salguero‐Gómez, R., Jones, O. R., Archer, C. R., Bein, C., de Buhr, H., Farack, C., ... & Römer, G. (2016). COMADRE: a global database of animal demography. Journal of Animal Ecology, 85(2), 371-384.